Profound Effects of Combining Choline and Piracetam on Memory Enhancement and Cholinergic Function in Aged Rats1
RAYMOND T. BARTUS*t, REGINALD L. DEAN, III*, KATHLEEN A. SHERMANt, EITAN FRIEDMANt* AND BERNARD BEER*
*Department of CNS Research, Medical Research Division of American Cyanamid Company
Lederle Laboratories, Pearl River, NY 10965 and tDepartments of Psychiatry< and PharmacologyX, New York University School of Medicine
New York, NY 10016
Received 30 March 1981
BARTUS, R. T., R. L. DEAN, III, K. A. SHERMAN, E. FRIEDMAN AND B. BEER. Profound effects of combining choline and piracetam on memory enhancement and cholinergic function in aged rats. NEUROBIOL. AGING 2(2) 105-111, 1981.—In an attempt to gain some insight into possible approaches to reducing age-related memory disturbances, aged Fischer 344 rats were administered either vehicle, choline, piracetam or a combination of choline or piracetam. Animals in each group were tested behaviorally for retention of a one trial passive avoidance task, and biochemically to determine changes in choline and acetylcholine levels in hippocampus, cortex and striatum. Previous research has shown that rats of this strain suffer severe age-related deficits on this passive avoidance task and that memory disturbances are at least partially responsible. Those subjects given only choline (100 mg/kg) did not differ on the behavioral task from control animals administered vehicle. Rats given piracetam (100 mg/kg) performed slightly better than control rats (p<0.05), but rats given the piracetam/choline combination (100 mg/kg of each) exhibited retention scores several times better than those given piracetam alone. In a second study, it was shown that twice the dose of piracetam (200 mg/kg) or choline (200 mg/kg) alone, still did not enhance retention nearly as well as when piracetam and choline (100 mg/kg of each) were administered together. Further, repeated administration (1 week) of the piracetam/choline combination was superior to acute injections. Regional determinations of choline and acetylcholine revealed interesting differences between treatments and brain area. Although choline administration raised choline content about 50% in striatum and cortex, changes in acetylcholine levels were much more subtle (only 6-10%). No significant changes following choline administration were observed in the hippocampus. However, piracetam alone markedly increased choline content in hippocampus (88%) and tended to decrease acetylcholine levels (19%). No measurable changes in striatum or cortex were observed following piracetam administration. The combination of choline and piracetam did not potentiate the effects seen with either drug alone, and in certain cases the effects were much less pronounced under the drug combination. These data were discussed as they relate to possible effects of choline and piracetam on cholinergic transmission and other neuronal function, and how these effects may reduce specific memory disturbances in aged subjects. The results of these studies demonstrate that the effects of combining choline and piracetam are quite different than those obtained with either drug alone and support the notion that in order to achieve substantial efficacy in aged subjects it may be necessary to reduce multiple, interactive neurochemica) dysfunctions in the brain, or affect activity in more than one parameter of a deficient metabolic pathway.
Choline Piracetam Memory enhancement Cholinergic function Aged rats
DURING the last several years a number of studies have indicated that age-related dysfunctions in central cholinergic mechanisms play an important role in the memory loss observed in elderly humans and patients suffering from senile dementia (i.e., [3, 15, 34, 35]). This hypothesis has prompted interest in therapeutically enhancing cholinergic activity to reduce the memory loss. Independent biochemical evidence has demonstrated that under certain conditions, increasing the availability of choline or lecithin (precursors for acetylcholine) may enhance cholinergic transmission [12,25], Although some evidence has been published that increasing amounts of choline or lecithin can improve performance on
certain behavioral tasks [2, 7, 13, 39], attempts to improve memory performance in aged humans (for review see [5]) or animals have not succeeded [8]. It has been suggested that one reason for this failure in geriatric subjects may be that the aged brain is unable to incorporate extra amounts of choline into acetylcholine, as reportedly occurs in younger brain tissue [5,6]. It is also possible that it may be necessary to improve other factors in aged brains before substantial increases in presynaptic cholinergic effects are obtained with precursor loading. For example, although normal cholinergic activity is dependent upon intact oxidative metabolism, it is known that several parameters which reflect energy produc-
‘Portions of these data were previously reported at the 12th Collegium Internationale Neuro-Psychopharmacologicum (CINP), Goteborg, Sweden, June, 1980 and the 10th Annua] Meeting of the Society for Neuroscience, Cincinnati, OH, November, 1980.
tion are decreased in the aged central nervous system [33, 38, 40]. Further, although the conversion of choline into acetylcholine occurs more readily under conditions of increased neuronal activity, recent circumstantial evidence suggests the activity of certain cholinergic pathways may be reduced in aged subjects [37]. Thus, either of these (or similar) factors could contribute to a situation in the aged brain that would prohibit extra choline from being effectively utilized for the synthesis of additional acetylcholine, and in turn, explain the negative results obtained with precursor studies in aged animals and humans.
One way to attempt to compensate for these possible age-related deficits would be to administer abundant amounts of choline while simultaneously giving a drug which might correct other critical age-related neuronal deficiencies. Although no drug yet exists that is recognized as effective in correcting neuronal function in aged brain, one drug which is beginning to attract interest for its biochemical and pharmacological properties is piracetam. Piracetam (2-pyrrolidine acetamide) is a relatively novel drug marketed in Europe for geriatric indications [18]. It has been reported to improve learning and memory in both animals and humans (for review see [17,18]) although its efficacy in aged subjects is not robust and is still controversial [1, 16, 23, 31]. Several lines of pharmacological evidence indicate that piracetam enables the central nervous system to function more effectively under hypoxic conditions [34], and that behavior is improved in oxygen deprived [36] or aged animals [11,41]. Neurochemical determinations suggest piracetam may facilitate conversion of ADP to ATP [22, 28, 34]. Other tests indicate piracetam also enhances intercerebral neuronal activity [10,18], and may deplete hippocampal tissue acetylcholine levels [44], Given this profile, it seems possible that piracetam might be able to reduce certain metabolic deficiencies in the aged brain which could normally contribute to the lack of significant effects observed with choline loading. This possibility was tested with aged Fischer 344 rats administered with choline, piracetam, or combinations of each and measuring effects on retention of a one-trial passive avoidance task.
Aged Fischer 344 rats had previously been shown to suffer severe impairments on this task as a natural consequence of aging [30]. Control studies suggested that a major source of this impairment involves a loss of memory for the learned event. For example, possible differences in motor activity or shock threshold cannot explain the age-related differences in the test day [30]. Further, evaluations of performance after various retention intervals demonstrated that the performance of the aged rats was comparable to the young rats when tested within an hour after training, but decreased sharply, exhibiting severe deficits within 4 hours after training [30]. These findings strongly suggest a memory-related phenomenon is involved with this age deficit. Because many operational characteristics of the task and the deficit are conceptually similar to the memory losses observed in other aged mammalian species, including mice [14], old world monkeys [8], new world monkeys [6] and humans [9], this procedure was used as a model system in the present study to evaluate the effects of choline and piracetam in aged subjects.
EXPERIMENT 1 METHOD
Subjects
Fifty-nine male Fischer 344 rats were used, ranging in age
between 20 to 29 months old. The animals were housed individually with food (Fischer Lab Chow«) and water available ad lib. The animal colony room was maintained under a 12 hour lightrdark cycle with a temperature of 22 ± 2 °C and a relative humidity of 40 to 45%.
Behavioral Procedures
A single-trial, step-through passive avoidance task was used. Because the apparatus and procedure have been reported previously [30], only a brief description will be presented here. The apparatus consisted of a two-chamber box which was separated by a guillotine door. On the initial training day, each subject was placed in the front, illuminated chamber with the guillotine door in the lowered (closed) position. After a 3 second orientation period, the door was raised, allowing the rat to freely explore the apparatus, whereupon it soon entered the darker, rear chamber. Immediately upon entering the rear chamber the guillotine door was lowered, preventing the rat from escaping back to the illuminated chamber, and a 1 mA electric shock was delivered to the feet through the floor grids for 3 seconds. Following this adversive experience, the rat was removed from the apparatus and returned to its home cage to await retention testing 24 hours later. Testing was accomplished by returning the rat to the apparatus and placing it in the front chamber as had been done on the previous day. The latency to re-enter the rear chamber (in which the rat had previously been shocked) was recorded. Control tests have indicated that the higher the latency to re-enter the rear chamber, the greater the apparent retention of the prior footshock.
Drug Treatment
Four separate groups were tested: (1) control, (2) piracetam, (3) choline, and (4) a piracetam/choline combination. Testing was conducted in a counterbalanced manner, with the various ages of the subjects equally distributed across the four experimental groups. All treatments were administered chronically for 1 week in the animals drinking water at a concentration calculated to provide a dose of 100 mg/kg/day of each drug. The dose for piracetam was chosen on the basis of published pharmacological data that this would produce maximal effects in rats, and for choline that this dose exceeded that which would be expected to reliably raise choline blood levels by the oral route. The control group was given tap water.
Fluid intake was measured daily and no significant difference was found between any drug group. Earlier tests indicated that the mean daily fluid intake for our colony was 35 ml per rat.
Thirty minutes prior to passive avoidance training and retention testing, each subject was injected (IP) with a supplemental dose of either saline, choline (60 mg/kg), piracetam (100 mg/kg) or the combination, according to the experimental group to which they belonged. This was done to assure optimal blood and brain levels of each drug treatment, and these single doses were again selected on the basis of available literature relevant to intraperitoneal administration [14,24],
RESULTS
The results of this study indicated that while no differences in latency to enter the rear chamber existed on the training day, large differences between treatments were ob-
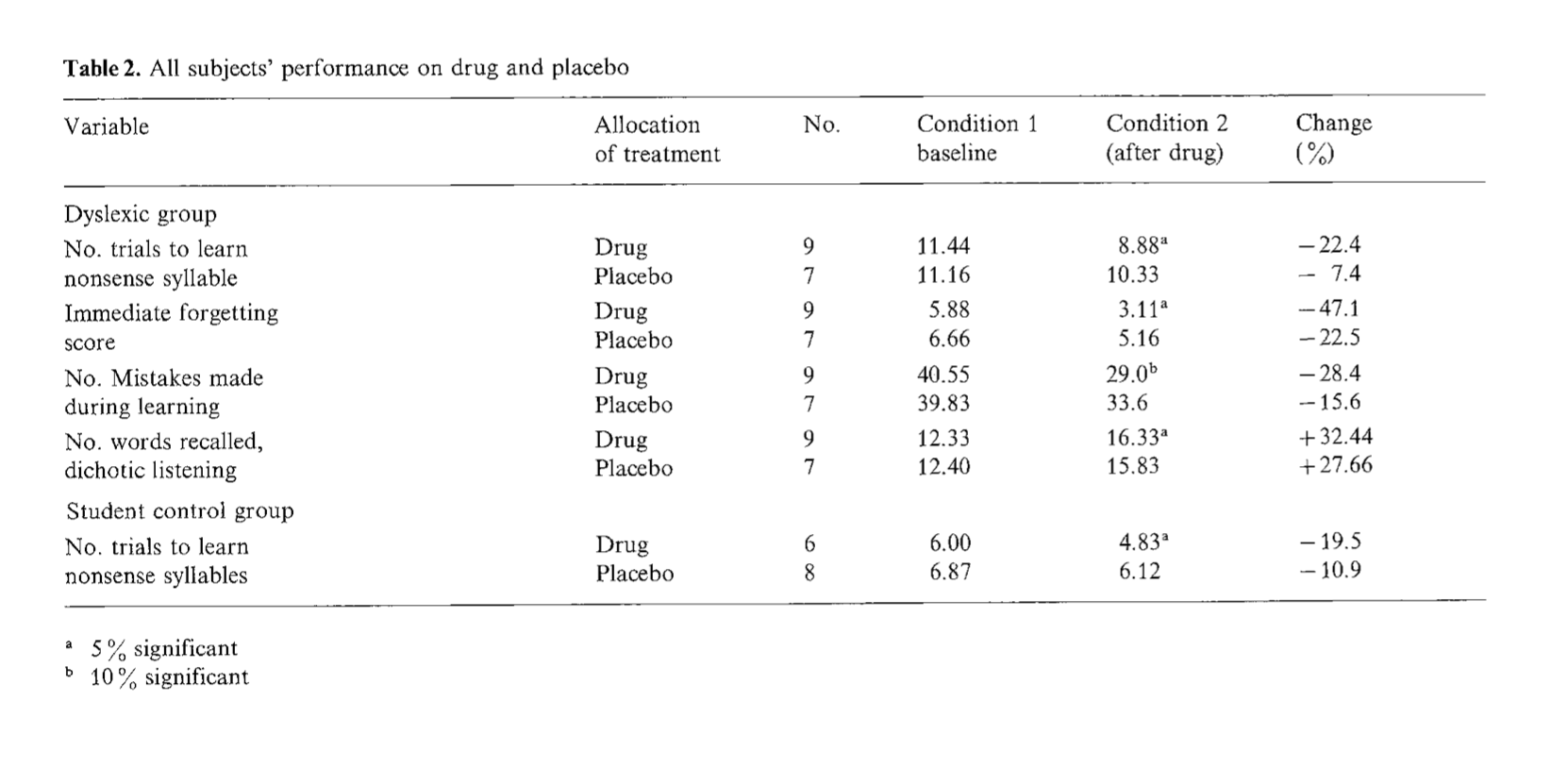
CHOLINERGIC FUNCTION AND MEMORY
served on the test day (Fig. 1). These differences were confirmed by an analysis of variance, F(3,39)=22.2, p<0.0001. The retention scores in the control group were quite consistent with the deficits previously obtained in this task situation with this strain of aged rat [30]. Individual comparisons of the treatments with the control group indicated that while choline did not significantly enhance retention latencies, f(20)= 1.964, p>0.05, piracetam did produce a subtle, but statistically reliable improvement on retention scores, f(20)=3.7, p<0.005. However, the retention scores for the piracetam/choline group was much more robust, f(19)=6.22,
p<0.0001.
REPLICATION
In order to replicate and extend these findings another experiment was performed. Its major purposes were to determine whether the improvement under the piracetam/ choline combination was simply due to additive effects of the individual treatments (or alternatively represent some synergistic effect), and to evaluate the relative contributions of the acute vs one week chronic treatments.
METHOD
Subjects
Forty additional naive male Fischer 344 rats were used for this replication. All animals were 20 months old at the initiation of this study and were housed in the same manner reported for Experiment 1.
The same behavioral test apparatus and procedure was used as in the first experiment. Five different drug treatments were evaluated, in addition to a saline control group, as shown in Table 1. The first three treatments were tested to help determine the relative contribution of the acute vs chronic treatments. They consisted of: (a) single acute injections of the drug combination (100 mg/kg of each) given before training and testing, (b) a 1 week dosing regimen of the combination (100 mg/kg of each), with a saline control injection given before training and testing, and (c) the combined 1 week combination of choline and piracetam with a supplemental injection (similar to that of the first experiment). Two additional treatments were tested, one of which gave twice
FIG. 1. Effects of saline, choline (100 mg/kg), piracetam (100 mg/kg), or choline and piracetam (C+P; 100 mg/kg each), administered to aged rats (23 to 29 months old) for a duration of 1 week prior to training and testing on a single-trial passive avoidance task. Previous research demonstrated that rats of this age suffer a severe impairment when tested for retention on the task, and that a deterioration in memory for the training trial contributes significantly to this deficit. Note synergistic-like effect on retention when choline and piracetam were given concurrently.
the dose of choline (200 mg/kg) for 1 week with supplemental injections before training and testing, and the other giving twice the dose of piracetam (200 mg/kg) according to the same protocol. These two groups were tested to help determine whether the robust positive effects obtained with the piracetam/choline combination in the first experiment could be obtained by simply giving greater amounts of either piracetam of choline alone.
RESULTS
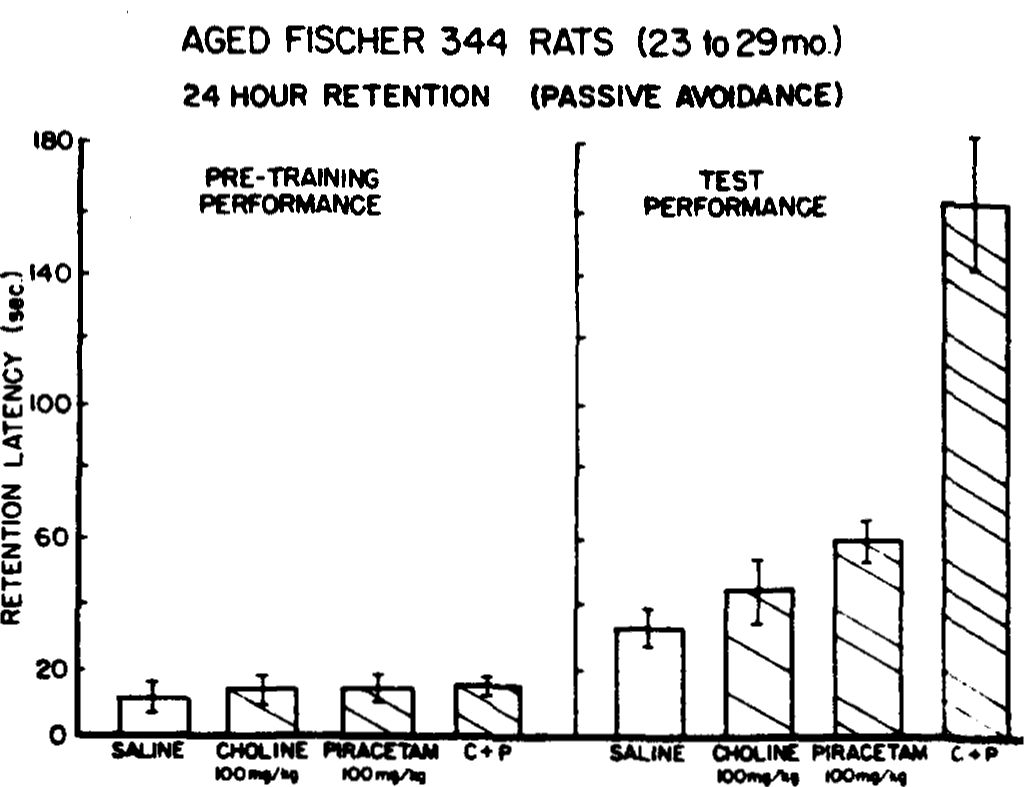
AGED FISCHER 344 RATS (20 mo.) 24 HOUR RETENTION (PASSIVE AVOIDANCE )
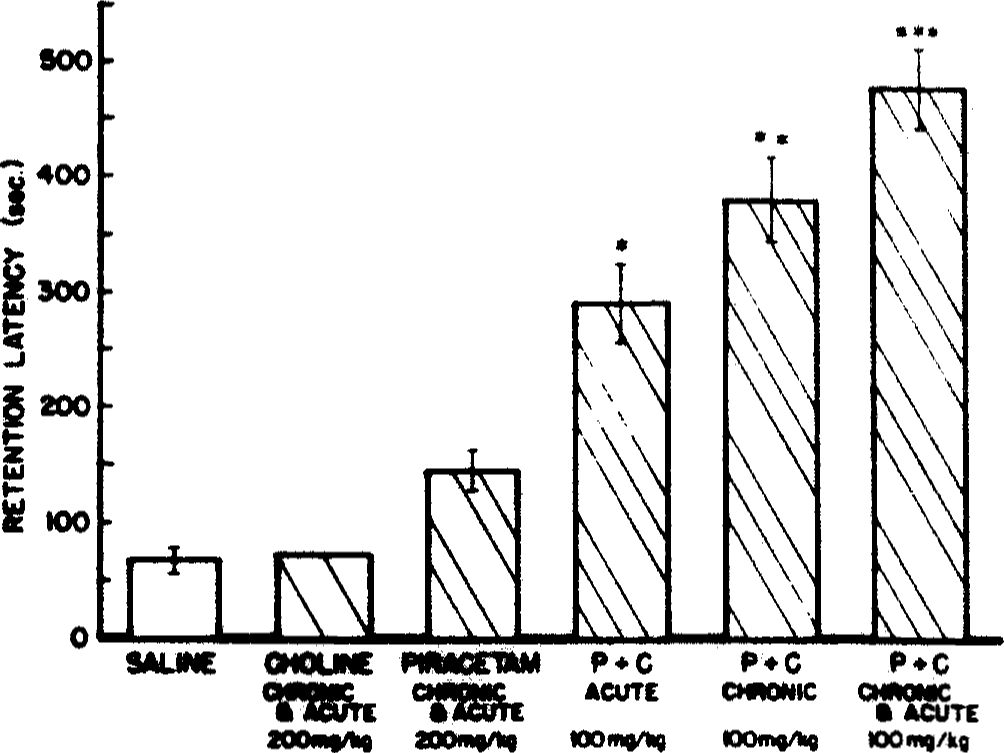
FIG. 2. Effects of saline, choline (200 mg/kg), piracetam (200 mg/kg) or piracetam and choline (P+C; 100 mg/kg each), administered to aged rats (20 months old) prior to training and testing on a single-trial passive avoidance task. Acute refers to single injections 30 min prior to training and testing, whereas chronic refers to administration for 1 week (in their water) prior to training and testing. (*p<0.05;
<0.005; ***p<0.001, according to Sheffe.)
The results of this study are illustrated in Fig. 2 and once again demonstrate substantial differences between treatments, F(4,28)=16.48,p<0.0001. (Because only one subject on the choline treatment survived the injections, this group could not be included in the statistical analysis; clearly, however, performance under this treatment was not improved and higher doses could not be given.) Individual comparisons between treatments were made using the Sheffe method of multiple comparisons [20]. According to this conservative analysis, although piracetam (at 200 mg/kg) produced a clear trend toward improvement, this difference was not statistically reliable when compared to control, F(4,28)=1.35, p>0.05. On the other hand, the combination of piracetam and choline again produced reliable improvement in retention. The acute dose (Treatment 4, Table 1) produced the least robust effect when compared with control, F(4,28)=3.57, p<0.05, with greater and more reliable effects obtained in the chronic only treatment (Treatment 5), F(4,28)=4.99, p<0.005. Finally, the most dramatic effects were observed under the chronic plus acute dosing regimen (Treatment 6), F(4,26)=6.9, p<0.001. In fact this treatment differed significantly from all others except the chronic only combination (Treatment 5). Thus, these data demonstrate that twice the dose of piracetam (or choline) alone did not produce as reliable or robust effects as when the two agents were administered simultaneously. Further, the longer the combination was administered the greater the apparent effects obtained.
BIOCHEMICAL DETERMINATIONS
METHOD
Following behavioral testing, rats from appropriate treatment groups were randomly selected for regional determinations of choline and acetylcholine. These animals had been part of the control, choline (100 mg/kg), piracetam (100 mg/kg) or piracetam/choline (100 mg/kg) treatments and were maintained on these regimens for a total of 14 days before being sacrificed. Thirty minutes before sacrifice, the animals were given supplemental injections of either saline, piracetam (100 mg/kg), choline (60 mg/kg) or the combination of each, as had been done prior to behavioral testing in Experiment 1.
The rats were sacrificed by microwave irradiation (2.5 kW, 2.45 MHz) focused on the head for 4 sec using a modified Litton microwave oven (Medical Engineering Consultants, Lexington, MA). Brain areas were dissected according to Glowinski and Iversen [21] and homogenized in 0.4 N HCIO, for 20 sec (Polytron homogenizer, Brinkman) and spun for 20 min at 4 °C and 25,000 xG. The pH of the supernatant was adjusted to pH 4.0 with potassium acetate (7.5 N) and samples were processed for determination of acetylcholine (ACh) and choline (Ch) according to the gas chromatographic method of Jenden and Hanin [27] using a Sigma 3 gas chromatograph equipped with a rubidium bead detector. The concentrations of ACh and Ch were calculated on the basis of the peak height ratios of their corresponding demethylated analogs to the peak height of dimethyl-aminomethylbutyrate, the demethylated analog of the internal standard, butyrocholine.
RESULTS
The effect of the drug treatments on Ch and ACh content of hippocampus, striatum and cortex are shown in Table 2. Differences between the treatments and control group were assessed by non-parametric statistical analyses [32]. Following Ch administration, the brain Ch content was increased about 50% in all three regions, although the increase was significant only in cortex (Rank sums=66 vs 124, p =0.027) and striatum (Rank sums=45 vs 108, p =0.004). ACh content tended to increase subtly after Ch treatment in these two regions (Rank sums=60 vs 130, p<0.05; and 57 vs 96, p <0.089, respectively), but not in hippocampus (p>0.10). Piracetam alone resulted in a marked increase (88%; Rank sums=49 vs 87, p=0.001) in hippocampal Ch content, and a 19% reduction of ACh (Rank sums=104 vs 67, p =0.057) in this brain region. However, no reliable effects on Ch or ACh concentrations were observed in striatum or cortex after piracetam treatment (p>0.10). The combination of piracetam and Ch treatment did not potentiate the trends observed after the administration of either drug alone. In fact, the hippocampal Ch content was increased less after combination treatment than after either piracetam or Ch alone, and the trend toward ACh reduction was less pronounced after combined treatment.
GENERAL DISCUSSION
Several interesting findings have emerged from this series of studies. First, while choline was ineffective in improving retention, and piracetam was only marginal, the two agents together induced striking improvement on the task. Twice the dose of either piracetam or choline alone did not produce effects which even approached the combination, suggesting
that the behavioral changes were due to some synergistic-like action of the two agents. The clear superiority of the combination therefore supports the proposal that in order to achieve substantial efficacy in aged subjects, it may be necessary to reduce multiple, interactive neurochemical dysfunctions or affect activity in more than one parameter of a deficient metabolic pathway [4],
The advantages of repeated administration and/or maintaining effective brain/blood levels in aged subjects for a period of at least several days seems apparent from the results of the second study (Fig. 2). These data in aged rats therefore agree with recent reports of memory facilitation in aged monkeys following chronic, but not acute administration of piracetam and similar pharmacological agents [4]. In fact, rarely have significant, positive effects in aged subjects been reported with acute doses of drugs. However, it still needs to be demonstrated that behavioral effects equally impressive to those reported here can be obtained in other task procedures, using other species, particularly humans. The robustness of the age-related passive avoidance deficit in rodents, the apparent involvement of CNS-related disturbances in the deficit, and the existence of certain operational similarities between this deficit and the primary memory loss observed in aged humans and non-human primates raises the possibility that the procedure used in the present studies may have some predictive value in geriatric research and that these positive effects may therefore generalize to other procedures and species. Certainly, the consistent lack of effects seen under choline, and the marginal effects obtained with piracetam are consistent with much of the other animal and human geriatric literature. Further, it is encouraging that a recent clinical trial based on these preliminary animal data found significant improvement in 3 of 10 mild to moderate Alzheimer’s patients following combined choline and piracetam treatment for 1 week [17].
In our initial investigation of the neurochemical mechanisms underlying the behavioral effects of piracetam and choline treatment, we have analyzed brain regional content of choline and acetylcholine. The results obtained in these
experiments suggest that the synergistic effect of choline plus piracetam combination is not simply due to an effect of piracetam on the uptake of Ch into the brain. The increase in regional Ch content was either the same or in fact, slightly less marked in rats receiving combination treatment than in those treated with either drug alone. After repeated piracetam treatment, a marked rise in Ch occurred specifically in the hippocampus. It is unclear at present whether this effect relates to an interaction of piracetam with phospholipid metabolism [43] or to a specific action on cholinergic mechanisms. It is also unclear why this effect was restricted to the hippocampus.
The effect of these drug regimens on regional ACh content were relatively small, and borderline in terms of statistical significance. Nevertheless, the trends observed here may provide interesting (albeit tentative) information regarding possible interactions of these treatments with cholinergic neurons. Repeated piracetam administration resulted in regionally specific alterations of ACh and Ch content. The reduction of hippocampal ACh content occurring in conjunction with an increase in Ch, could indicate either an inhibition of ACh synthesis, or alternatively, an increase in ACh release after piracetam. Given the pharmacological profile of piracetam, including the behavioral improvement observed in the present study, it is unlikely that piracetam inhibits ACh synthesis. The effect of chronic piracetam treatment on hippocampal ACh observed here in aged rats is consistent with changes recently reported to occur in young rats following acute piracetam, but differ in that a change in hippocampal Ch was not reported in the young rats after acute treatment [44],
The possibility that piracetam results in enhanced ACh release from septo-hippocampal neurons is especially intriguing in view of our recent observations suggesting that cholinergic neuronal activity and/or ACh release are impaired in hippocampus of aged rats (<20 months) [37]. Additional evidence of age-related impairment in cholinergic transmission in this brain region has been provided by elec-trophysiological analyses showing that the basal firing rate of
hippocampal pyramidal cells is reduced and the responsiveness of these neurons to ACh is diminished with age in rats [29], It is therefore suggested that one effect of piracetam may be to partially reverse an age-related deficit in hippocampal cholinergic transmission by increasing hippocampal neuronal activity and/or ACh release, and that this action may play a role in the behavioral improvement observed. The partial reversal of hippocampal ACh depletion after piracetam/choline combination may, at first, seem inconsistent with this hypothesis. However, it has previously been shown that when choline is administered in coiyunction with drugs known to enhance ACh release, such as atropine, the consequent decline in hippocampal ACh content may be partially antagonized [42]. Clearly, it will be necessary to utilize more direct measures of ACh release (than changes in ACh content) in future experiments in order to empirically test the relationship between effects of piracetam or piracetam/choline treatment on cholinergic transmission and on retention test performance. In addition, other possible mechanisms of the marked improvement of retention associated with piracetam plus choline may be considered. For example, recent evidence suggests that piracetam affects phospholipid metabolism [43] and the role of choline in phospholipid metabolism is well established [24]. These ef
fects could conceivably lead to a reduction in certain age-related deficits in post-synaptic receptor mechanisms or other membrane-related phenomena which may play an important role in the behavioral disturbances associated with aging [7].
Regardless of the particular neurochemical factors responsible for the behavioral changes observed here, the present study clearly shows that the neurochemical and be havioral effects of combining piracetam and choline are quite different than either agent alone. The robust interactive affects may represent a phenomenon or principle fundamental to the successful treatment of age-related behavioral dysfunctions.
ACKNOWLEDGEMENTS
The authors acknowledge the comments and constructive criticism of Drs. A. S. Lippa and L. R. Meyerson (Department of CNS Research, American Cyanamid), assistance with the statistical analyses by Drs. R. Rechtschaffner, T. Hoffman and S. Heft (Statistical Department, American Cyanamid) and the assistance of R. Sheppard (Department of CNS Research) in the preparation of this manuscript.
REFERENCES
1. Abuzzahab, F. S., G. E. Mervin, R. L. Zimmermann and M. C. Sherman. A double blind investigation of piracetam (Nootropil) vs. placebo in geriatric memory. Pharmakopsychiatrie 10: 49-56, 1977.
2. Barbeau, A. Lecithin in neurological disorders. New Engl. J. Med. 299: 200-201, 1978.
3. Bartus, R. T. Cholinergic drug effects on memory and cognition in animals. In: Aging in the 1980’s: Psychological Issues, edited by L. W. Poon. Washington, DC: American Psychological Association, 1980.
4. Bartus, R. T. and R. L. Dean. Age-related memory loss and drug therapy: Possible directions based on animal models. In: Aging, Vol. 17: Brain Neurotransmitters and Receptors in Aging and Age-Related Disorders, edited by S. J. Enna, T. Samorajski and B. Beer. New York: Raven Press, 1981.
5. Bartus, R. T. and R. L. Dean. The cholinergic hypothesis of impaired memory in the aged. Nutritional and pharmacological implications. In: Aging, Vol. 18: Nutrition and Aging, edited by D. Harman and J. M. Ordy. New York: Raven Press, in press.
6. Bartus, R. T., R. L. Dean and B. Beer. Memory deficits in aged Cebus monkeys and facilitation with central cholinomimetics. Neurobiol. Aging 1: 145-152, 1980.
7. Bartus, R. T., R. L. Dean, J. A. Goas and A. S. Lippa. Age-related changes in passive avoidance retention: Modulation with dietary choline. Science 209: 301-303, 1980.
8. Bartus, R. T., D. Fleming and H. R. Johnson. Aging in the rhesus monkey: Effects on short-term memory. J. Geront. 33: 858-871, 1978.
9. Botwinick, J. Behavioral processes. In: Aging, Vol. 2: Genesis and Treatment of Psychological Disorders in the Elderly, edited by S. Gershon and A. Raskin. New York: Raven Press, 1975.
10. Buresova, D. and J. Bures. Mechanisms of interhemispheric transfer of visual information in rats. Acta Neurobiol. exp. 33: 673-688, 1973.
11. Bumotte, R. E., J. G. Gobert and J. J. Temmerman. Piracetam (2-pyrrolidine acetamide) induced modifications of the brain polyribosome pattern in ageing rats. Biochem. Pharmac. 22: 811-814, 1973.
12. Cohen, E. L. and R. J. Wurtman. Brain acetylcholine: Increases after systemic choline administration. Life Sci. 16: 1095-1102,
1975.
13. Davis, K. L., L. E. Hollister, J. D. Barchas and P. A. Berger. Choline in tardive dyskinesia and Huntington’s disease. Life Sci. 19: 1507-1516, 1976.
14. Dean, R. L., J. A. Goas, B. Regan, J. Scozzafava and R. T. Bartus. Age-related differences in the lifespan of the C57BL/6J mouse. Exps Aging res. 1981, in press.
15. Drachman, D. and J. Leavitt. Human memory and the cholinergic system: A relationship to aging? Archs Neurol. 30: 113-121, 1974.
16. Ferris, S. H. and B. Reisberg. Clinical trials: Neuropeptides and nootropics. Neurobiol. Aging 2: 71, 1981.
17. Friedman, E., K. A. Sherman, S. H. Ferris, B. Reisberg, R. T. Bartus and M. K. Schneck. Clinical response to choline plus piracetam in senile dementia: Relation to red-cell choline levels. New Engl. J. Med. 304: 1490-1491, 1981.
18. Giurgea, C. Piracetam: Nootropic pharmacology of neurointcg-rative activity. Curr. devs Psychopharmac. 3: 221-273, 1976.
19. Giurgea, C. and M. Salama. Nootropic drugs. Prog. Neuro-Psychopharmac. 1: 235-247, 1977.
20. Glass, G. V. and J. C. Stanley. Statistical Methods in Education and Psychology. Englewood Cliffs, NJ: Prentice-Hall, 1970.
21. Glowinski, J. and L. L. lversen. Regional studies of catecholamines in rat brain. I. The disposition of (*H)-norepinephrine, (*H)-dopamine and (’HpDOPA in various regions of the brain. J. Neurochem. 13: 655-669, 1966.
22. Gobert, J. G. and J. J. Temmerman. Piracetam induced modification of the brain polyribosome content in ageing rats. In: Al-tern, edited by D. Platt and F. K. Schattaner. New York: Springer-Verlag, 1973.
23. Gustafson, L., J. Risberg, M. Johanson, M. Fransson and V. A. Maximilian. Effects of piracetam on regional cerebral blood flow and mental function in patients with organic dementia. Psychopharmacology 56: 115-117, 1978.
24. Haubrich, D. R., N. H. Gerber and A. B. Pflueger. Choline availability and the synthesis of acetylcholine. In: Nutrition and the Brain, Vol. 5: Choline and Lecithin in Brain Disorders, edited by A. Barbeau, J. H. Growdon and R. J. Wurtman. New York: Raven Press, 1979.
25. Haubrich, D. R„ P. F. L. Wang, D. E. Clody and P. W. Wedek-ing. Increases in rat brain acetylcholine induced by choline and deanol. Life Sci. 17: 975-980, 1975.
26. Hirsch, M. J. and R. J. Wurtman. Lecithin consumption elevates acetylcholine concentrations in rat brain and adrenal gland. Science 202: 223-224, 1978.
27. Jenden, D. J. and I. Hanin. Gas chromatographic microestimation of choline and acetylcholine after N-demethylation of sodium benzothiolate. In: Choline and Acetylcholine: Handbook of Chemical Assay Methods, edited by I. Hanin. New York: Raven Press, 1974, pp. 135-150.
28. Kabes, J., L. Erban, L. Hanzlicek and V. Skonida. Biological correlates of piracetam: Clinical effects in psychotic patients. J. ini. Med. Res. 7: 277-284, 1979.
29. Lippa, A. S., D. J. Critchett, F. Ehlert, H. I. Yamamura, S. J. Enna and R. T. Bartus. Age-related alterations in neurotrans-mitter receptors: An electrophysiological and biochemical analysis. Neurobiol. Aging 2: 3-8, 1981.
30. Lippa, A. S., R. W. Pelham, B. Beer, D. J. Critchett, R. L. Dean and R. T. Bartus. Brain cholinergic function and memory in aged rats. Neurobiol. Aging 1: 10-16, 1980.
31. Lloyd-Evans, S., J. C. Brocklehurst and M. K. Palmer. Piracetam in chronic brain failure. Curt. med. Res. Opin. 6: 351-357, 1979.
32. Mann, H. B. and D. R. Whitney. On a test of whether one or two random variables is ptochastically larger than the other. Ann. math. Statist. 18: 50-60, 1947.
33. Meier-Ruge, W., O. Hunziker, P. I. Wangoff, K. Reichmeier and P. Sandoz. Alterations of morphological and neurochemical parameters of the brain due to normal aging. In: Developments in Neuroscience, Vol. 3: Senile Dementia: A Biochemical Approach, edited by K. Nandy. New York: Elsevier-North Holland, 1978.
34. Nickolson, V. J. and O. L. Wolthuis. Effect of the acquisitionenhancing drug piracetam on rat cerebral energy metabolism: Comparison with naftidrofuryl and methamphetamine. Biochem. Pharmac. 25: 2241-2244, 1976.
35. Perry, E. K., B. E. Tomlinson, G. Blessed, K. Bergmann, P. H. Gibson and R. H. Perry. Correlation of cholinergic abnormalities with senile plaques and mental test scores in senile dementia. Br. med. J. 2: 1457-1459, 1978.
36. Sara, S. J. and D. Lefevre. Hypoxia-induced amnesia in one-trial learning and pharmacological protection by piracetam. Psychopharmacology 25: 32-40, 1972.
37. Sherman, K. A., J. E. Kuster, R. L. Dean, R. T. Bartus and E. Friedman. Presynaptic cholinergic mechanisms in brain of aged rats with memory impairments. Neurobiol. Aging 2: 99-104, 1981.
38. Sims, N. R., C. C. T. Smith, A. N. Davison, D. M. Bowen, R. H. A. Flack, J. S. Snowden and D. Neary. Glucose metabolism and acetylcholine synthesis in relation to neuronal activity in Alzheimer’s disease. Lancet 1: 333-336, 1980.
39. Sitaram, J., H. Weingartner, E. D. Caine and J. C. Gillin. Choline: Selective enhancement of serial learning and encoding of low imagery words in man. Life Sci. 22: 1555-1560, 1978.
40. Sylvia, A. L. and M. Rosenthal. Effects of age on brain oxidative metabolism in vivo. Brain Res. 165: 235-248, 1979.
41. Valzelli, L., S. Bemasconi and A. Sala. Piracetam activity may differ according to the age of the recipient mouse. Int. Phar-macopsychiat. 15: 150-156, 1980.
42. Wecker, L., W. D. Dettbarn and D. E. Schmidt. Choline administrative modification of the central actions of atropine. Science 199: 86-87, 1978.
43. Woelk, H. Effects of piracetam on the incorporation of “P into the phospholipids of neurons and glial cells isolated from rabbit cerebral cortex. Pharmakopsychiatrie 12: 251-256, 1979.
44. Wurtman, R. J., S. G. Magil and D. K. Reinstein. Piracetam diminishes hippocampal acetylcholine levels in rats. Life Sci. 28: 1091-1093, 1981.