TOWARDS PHARMACOLOGY INTEGRATIVE ACTIVITY OF THE MIND
I. — INTRODUCTION
The term “integrative activity” was proposed at the beginning of the century by Sherrington, to define the central nervous processes that make starting from the basic reflexes one elaborates a voluntary movement. The integrative concept had been anticipated even before, by Hughlings Jackson who had written: “the simplest spinal reflex ‘thinks’ so to speak in movement and not in muscles “(after Sherrington, 1931). These reflexes elementary are “integrated” through a central mechanism.
Although initially considered at the spinal level, the notion of activity integrative was quickly used by the Sherringtonian school itself, in the physiological interpretation of the higher nervous levels.
Since then, research on brain function has taken place in two different ways, although complementary:
a) An analytical direction that uses histological methods and electrophysiological studies and seeks to establish the structural and functional details neurons and their interrelationships.
b) A synthetic direction that preferably uses methods behavioral and studies the brain as an organ of control of all the activity of the individual and the interaction with his environment. This is the direction initiated, also at the beginning of the century, by Pavlov and his school.
It does not seem possible at the present time to carry out a comprehensive study of of the brain, which would take into account all the knowledge lytic, anatomo-physiological available. Nevertheless, it is in this
direction that the neurological sciences are progressing, that is, toward a analytico-synthetic entanglement. This is how we see more and more research using the most advanced neurophysiological methods refined in awake subjects and even in free behavior. It will be recalled that the pioneers of this trend were Durup and Fessard (1935), Gastaut (1957), Livanov (1945), Delgado (1954), Roy John (1960) and Fessard (1960).
In what follows, we will introduce – based mostly on our own work – a new pharmacological approach to the problem of the integrative activity of the brain.
Not pretending to have a general view of the problem, but in order to situate it in the current context, we will first consider some neurophysiological and neuropharmacological pemisses.
II — NEUROPHYSIOLOGICAL PREMISSES AND NEUROPHARMACOLOGY
NEUROPHYSIOLOGY
The state of consciousness can go from wakefulness to sleep deep and coma. It is the expression of a certain cortical tone (Bremer, 1936) moderated by sensory, reticular, rhinencephalic afferences etc., which in turn modulates the periphery.
ROLE OF TELENCEPHALON IN THE INTEGRATIVE ACTIVITY OF C.N.S.
functional particularities still under study (Eccles, 1966a), the neurophysiology tells us that the cerebral cortex is composed of nerve cells that essentially resemble nerve cells lower floors of the nevraxe. The same goes for glial cells whose specific role in the integrative activities of the brain is postulated for a long time (Galambos, 1961). What differentiates the cerebral cortex other levels of the brain is its extreme complexity in the neuronal organization.
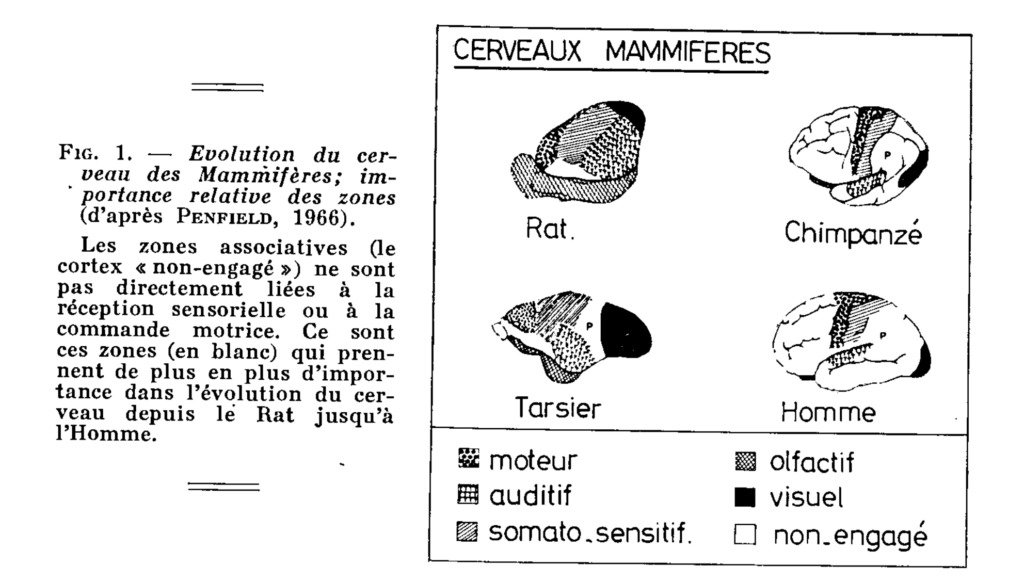
Evolution of the brain of mammals; relative importance of areas (Penfield 1966)
Associative areas (the “un-committed” cortex) are not directly related to sensory perception or motor control. It is these areas (in white) that gain more and more importance in the evolution of the brain from the rat to man.
This corresponds, moreover, to the physiological conclusion of Moruzzi (1966) that the percentage of cerebral activity which is directly related to perception is extremely small.
We thus return to Sherrington’s admirable intuition, which had said that “the essential character of the neuronal mechanism of the brain lies in its integrative power” (quoted from Eccles, 1966).
NEUROPHARMACOLOGY
From a neuropharmacological point of view, numerous investigations have shown the effect of chemical substances on learning and natural storage (McGauch and Petrinovitch, 1965).
It is not the purpose of this work to review them. We can only attempt a generalization that includes many exceptions, admitting that most of these drugs indirectly influence the integrative activities of the brain, by interventions on the processes of attention, perception or motivation. Thus, anticholinergics, barbiturates and depressives in general reduce the efficiency of learning and retention. On the contrary, anticholinesterases, stimulants or convulsants at subconvulsive doses, facilitate these same processes.
It should also be noted, with Roy John (1967) that substances such as strychnine, picrotoxin, nicotine, pentylene tetrazol, physostigmine, caffeine, amphetamine, and others that facilitate learning, are all CNS stimulants, but possess mechanisms of action well differentiated.
For example, it has been demonstrated that some stimulants accelerate the synthesis of RNAs in the brain (strychnine, tricyanoaminopropene, pemoline, diphenylhydantoine and procaine amide), while others, such as amphetamine, do not have this effect (Carlini). and Carlini 1965, Essman 1966, Glasky and Simon 1966, Gordon et al., 1968, Lima et al., 1966, Weissman 1967).
With the help of an example, we will try to demonstrate that this possibility has become a reality which opens, it seems to us, a new field of pharmacological news. This is the pharmacology of Piracetam, of which we will present here mainly the aspects related to the integrative activity of the brain.
Most of these aspects have already been the subject of specialized publications (to which we refer the reader every time), but not that of an interpretative synthesis whose present work is its first attempt.
III. – PIRACETAM AND THE TELENCEPHALIC HYPOTHESIS
THE “UNUSUAL” NEUROPHARMA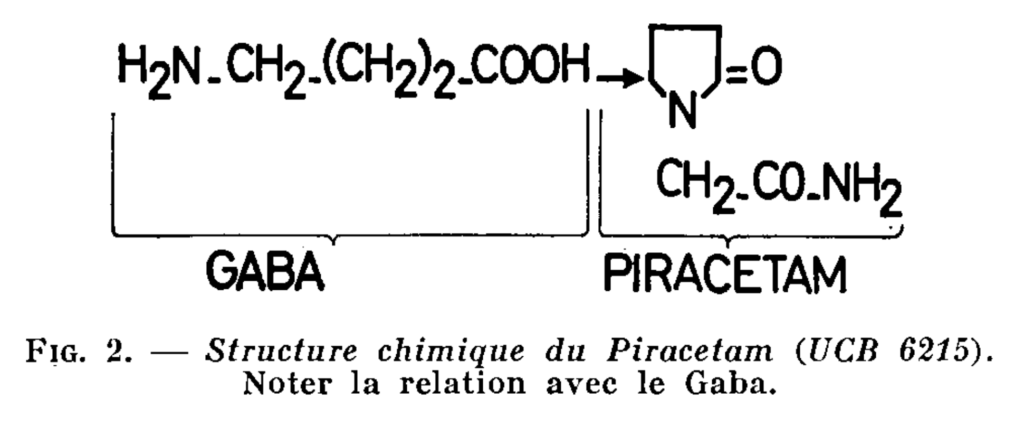 COLOGY OF PIRACETAM
COLOGY OF PIRACETAM
Piracetam is an original substance, chemically apparent in Gaba (see Fig. 2)
1 ° Inhibitory effect on central and vestibular nystagmus
Without being able to dwell here on the history of pharmacological research on Piracetam, which began in the years 1964-1965, it should be remembered that this product has been tried by us, for theoretical reasons explained elsewhere (Giurgea et al. , 1965), in the so-called “central” nystagmus test, that is to say obtained by electrical stimulation – the Rabbit – of the lateral knee body (Bergmann et al., 1959).
Piracetam administered intravenously or orally in a wide range of doses has been shown to be highly active, starting at 2 mg / kg, by inhibiting central nystagmus (Giurgea et al., 1967).
On a peripheral vestibular nystagmus, Piracetam has a similar effect in rabbits and in! Man (Giurgea and collaborators, 1967).
This effect of Piracetam was in itself a curiosity, because in these tests, only antihistamines and anticholinergics are active, but Piracetam is neither antihistamine nor anticholinergic, even if doses much higher than those which inhibit the excitability of the nystagmogenic structures.
2 ° Absence of behavioral, electroencephalographic or autonomic effects
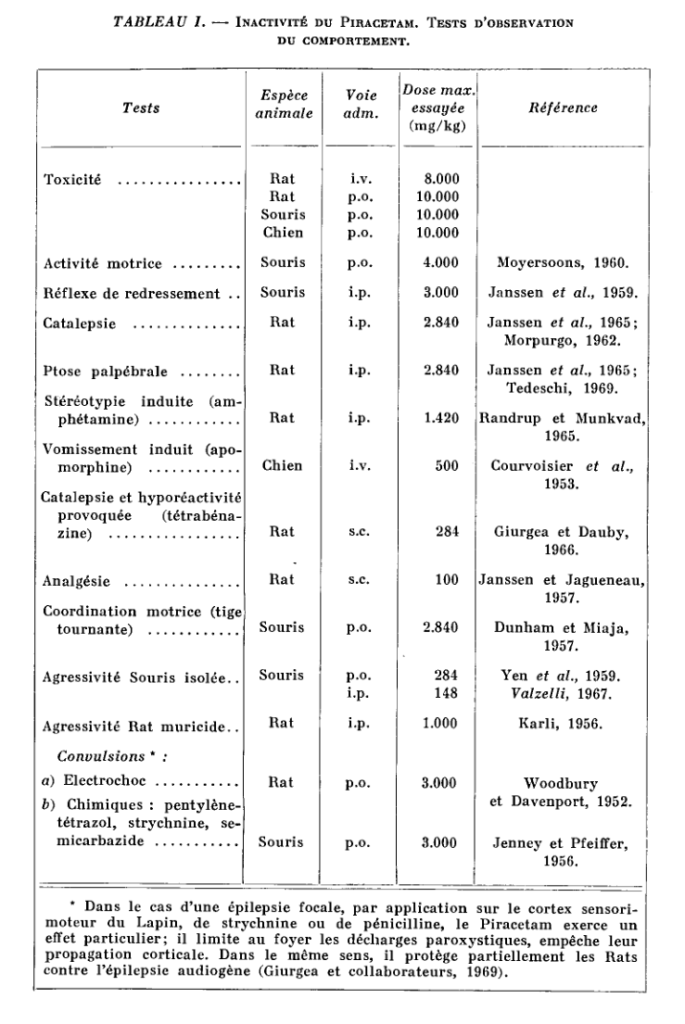
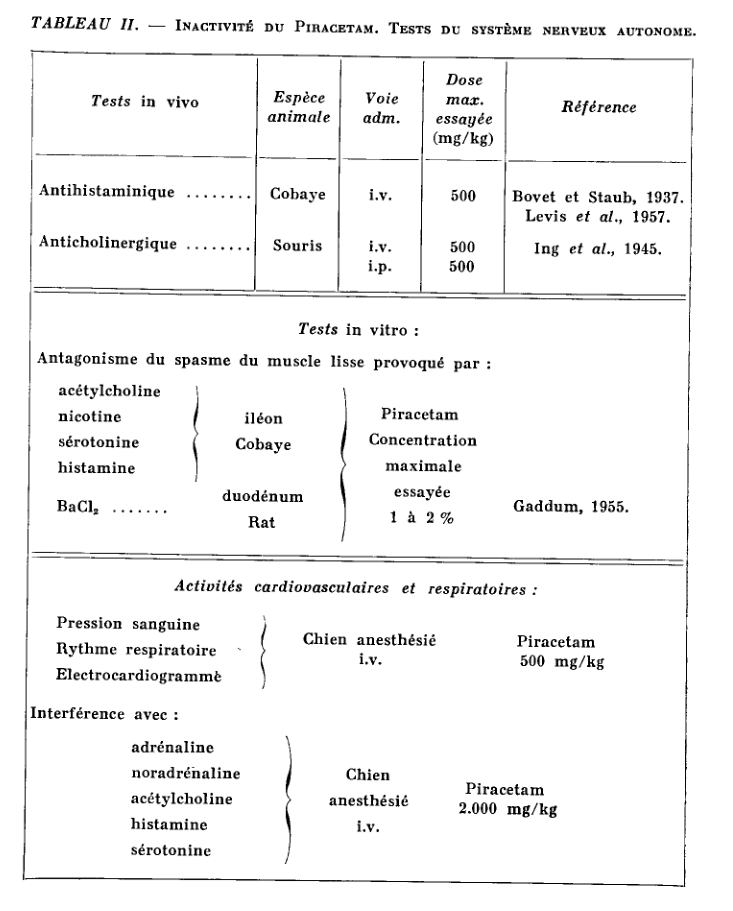
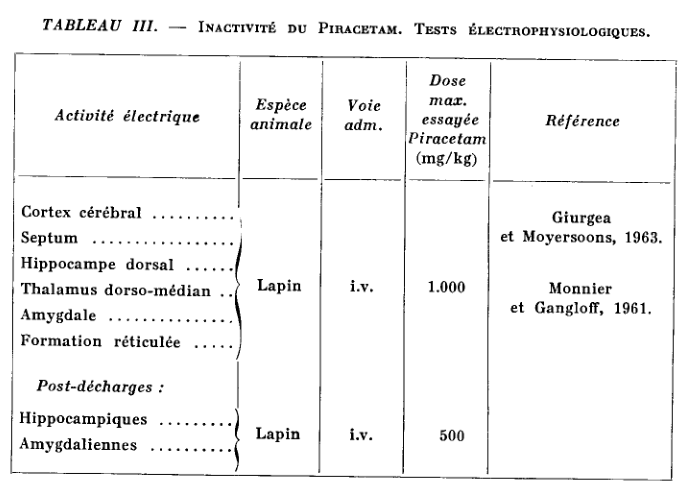
Piracetam not only has no antihistaminic or anticholinergic properties, but is inactive in most routine pharmacological tests (Giurgea et al., 1967).
Up to doses of a few g / kg intravenously or per os, in the mouse, rat, cat, dog and monkey, single or repeated administration, Piracetam is devoid of toxicity (in dogs, for example). for example, it has been possible to administer for one year the product by increasing the dose up to 10 g / kg / day) and does not produce behavioral, electroencephalographic or autonomic changes in the animal species studied.
The main tests in which Piracetam has been shown to be inactive in a wide range of doses are listed in Tables I, II and III.
In the case of focal epilepsy, by application to the sensorimotor cortex of Rabbit, strychnine or penicillin, Piracetam has a particular effect; it limits to the focus the paroxysmal discharges, prevents their cortical propagation. In the same sense, it partially protects rats against audiogenic epilepsy (Giurgea et al., 1969).
3 ° Protection on electroencephalographic pattern against hypoxia
The inhibitory effect of Piracetam on the central nystagmus as well as the absence of any foreseeable side effect, led us to propose clinical trials in the field of vertigo and in particular in those of central origin as in the case of cranial traumatises.
Given the encouraging results in this area (see page 147) and Taking into account the role of cerebral anoxia in the pathophysiology of cerebral trauma (Ingvar and Lassen, 1964) we hypothesized that Piracetam would protect against anoxic anoxia.
Therefore, we studied the effect of Piracetam on the evolution of electroencephalogram of rabbits subjected to deep anoxia by inhalation of nitrogen. It is known that under nitrogen, it produces a gradual deterioration of the electroencephalogram until “electrical silence” and that, if the animal is rapidly returned to the atmosphere, it recovers a normal electroencephalographic trace (Gastaut and Meyer, 1961, Pternitis and Passouant, 1961).
Under these conditions, we have found – comparing animals treated with saline solution to those treated with Piracetam – that Piracetam significantly increases the “resistance” cerebral (the time of appearance of electrical silence after the start of infusion of nitrogen in its cabin) but especially facilitates the “recuperation” cerebral (the time taken for the restoration of a normal electroencephalogram after readmission of the air). These experiments have been published elsewhere (Giurgea et al., 1970) and on this occasion we have shown that several analeptics such as benzedrine, methylphenidate, caffeine, theophylline and pentylenetetrazol not only do not exert a protective effect. but show rather toxic effects in this experimental procedure.
For illustration, we give a figure (Figure 3) showing the positive effect of Piracetam on cerebral recovery.We will see later that Piracetam has protection against hypoxia in other experimental situations as well.
4 ° Facilitation of memory and learning in normal animals or in a deficit state
(P125 Break)
(Pick up 149)
IV. – ATTEMPTING NOOTROPIC CONCEPT IN THE PHARMACOLOGY OF INTEGRATIVE BRAIN ACTIVITY